前脑中的少突胶质的竞争和胚胎谱系的后天消除
原文作者:
Nicoletta Kessaris1, Matthew Fogarty1, Palma Iannarelli1, Matthew Grist1, Michael Wegner2amp; William D Richardson1
单位:
1Wolfson Institute for Biomedical Research and Department of Biology, Universary College London, Gower Street, London WC1E 6BT,UK.2Institut fur Biochemie, Emil Fischer Zentrum, Fahrstrasse 17, 91054 Erlangen, Germany.
摘要:前脑少突细胞祖细胞(OLPs)的发育起源一直存在争议。我们现在通过在转基因小鼠的Crel - lox命运图谱中发现,第一个OLPs起源于腹侧前脑的内侧神经节隆起(MGE)和前眼内眼区(AEP)。从那里,他们植入了包括大脑皮层在内的整个胚胎端脑,然后再从侧面和/或尾部神经节的突出部分(LGE和CGE)加入第二波的OLPs。最后,第三波产生于产后皮质。当任何一个种群被白喉毒素的靶向表达所破坏时,其余的细胞就会被取代,而老鼠的生存和行为正常,它们是少突胶质细胞和髓磷脂的正常补充。因此,功能性冗余的OLPs种群在发育中的大脑中争夺空间。值得注意的是,胚胎期的MGE和AEP衍生的种群在出生后的生活中被消除了,这让人们对竞争的性质和目的产生了疑问。
关键词:少突胶质细胞;胚胎谱系;转基因小鼠
不同的神经元和神经胶质的亚类在发育的脊髓和大脑的脑室区域的不同部分相继产生。例如,脊髓运动神经元和少突胶质细胞(OLs,髓系形成细胞)来自于位于腹侧脑室区(运动神经元前体)的一个特殊领域内的前体(pMN),这是由转录因子Olig2的表达所定义的。(参考1-4)从那里,OLPs在分化成骨髓形成的OLs之前,全部通过脊髓迁移。之后,至少有一个额外的OLP来源出现在脊髓脊索,贡献10 - 15%的最终的OL 种群(参考5-7)。在pMN中,OLP生成依赖于信号分子 sonic hedgehog基因(Shh),而背源可能是独立Shh(参考5-9)。
在神经系统的前段(特别是前脑)中产生的OL不是很容易理解的。MGE的神经上皮和腹侧前脑的AEP表达了Shh及其受体的Patched(Ptc)和Olig2,这暗示着OLPs可能主要是在这些地区形成的(参考10-12)。实际上,由血小板源的生长因子receptor-a(Pdgfra)表达的迁移OLPs,在小鼠胚胎第12天(E12)后,从MGE和AEP的各个方向都可以看到,很明显是在E16周围进入了正在发育的大脑皮层(端脑背侧)(参考11,13)。在E16和出生(BE18)之间,皮质的OLPs的数量显著增加,但这是否反映了持续的向内迁移和腹侧衍生的OLPs或大脑皮层中新的OLPs来源的扩散,尚不清楚。双方都有一些证据。例如,大脑皮层的一些OLPs表达了Dlx1和Dlx2转录因子,表明它们起源于腹侧端脑,其中Dlx1和Dlx2是第一个激活的(参考14,15)。此外,在小鸡胚胎,少突胶质细胞的表达髓磷脂蛋白(Plp - Dm20)和O4抗原最初被限制在AEP(参考16),而chick - quail嫁接实验表明,所有的皮质醇都是在鸟中得到的(参考17)。相比之下,使用Emx1 - Cre转基因小鼠的细胞命运图谱显示,大脑皮层中的许多OLs是由基因组成的(参考18)。此外,逆转录病毒的命运图谱表明,皮质醇仍然是在出生后的时间内由位于皮质纹状边界的侧脑室的侧脑室的前体(SVZ)形成的(参考19,20)。解决这种不确定性是很重要的,因为不同发育来源的OLs,也许是通过不同的信号通路来指定的,在成熟的大脑中可能具有不同的性质和功能。
为了解决OL起源的问题,我们使用了转基因小鼠的Cre-lox方法,以追踪端脑中不同腹侧前体的发育情况。我们发现前脑中的OL谱系细胞的发育异常复杂和动态。在腹侧前脑的Nkx2.1 -表达前体中出现了早期的OLP生成波;这些最初出现在腹侧MGE的心室区,在E12.5周围出现AEP,随后广泛地迁移到端脑的所有部分,在E16后进入大脑皮层。一开始,他们是前脑里唯一的OLP,但很快他们对总OLP种群的贡献下降了。在出生后的第10天(P10),虽然在腹侧(subpallial)区域仍然是主要种群,但在皮质中很少有Nkx2.1-衍生的OLPs或OLs。取而代之的是,它们被其他的来自于LGE和CGE的阳性Gsh2-前体的OLPs细胞所取代,然后从内源性的Emx1 –阳性的皮质前体中获得。即使在腹侧前脑,接近其原始来源,Nkx2.1-衍生的OLPs和OLs逐渐消失并被其他种群取代,几乎完全从成人前脑消失。这些发现有助于整合以前的研究报告,包括腹侧背侧、背侧背侧或多侧鼻咽部(参考10-24)。
我们的研究结果提出了一个问题:MGE-AEP、LGE-CGE和皮质的不同的OL谱系是否具有功能上的专门化。为了解决这一问题,我们设计了一种基因消融策略,分别使用白喉毒素A片(DTA;参考25)。我们发现,当我们杀死任何一个种群时,其效果是良性的:邻近的种群扩散到空地上,恢复了正常的OLPs分布,小鼠正常发育并存活下来。因此,到目前为止,我们还没有发现前脑不同的OL谱系中功能异质性的证据。我们的数据表明,不同的区域亚种群在发育的大脑中争夺空间。这可能导致胚胎的MGE-AEP(Nkx2.1)-衍生 OLP种群的最终消亡。
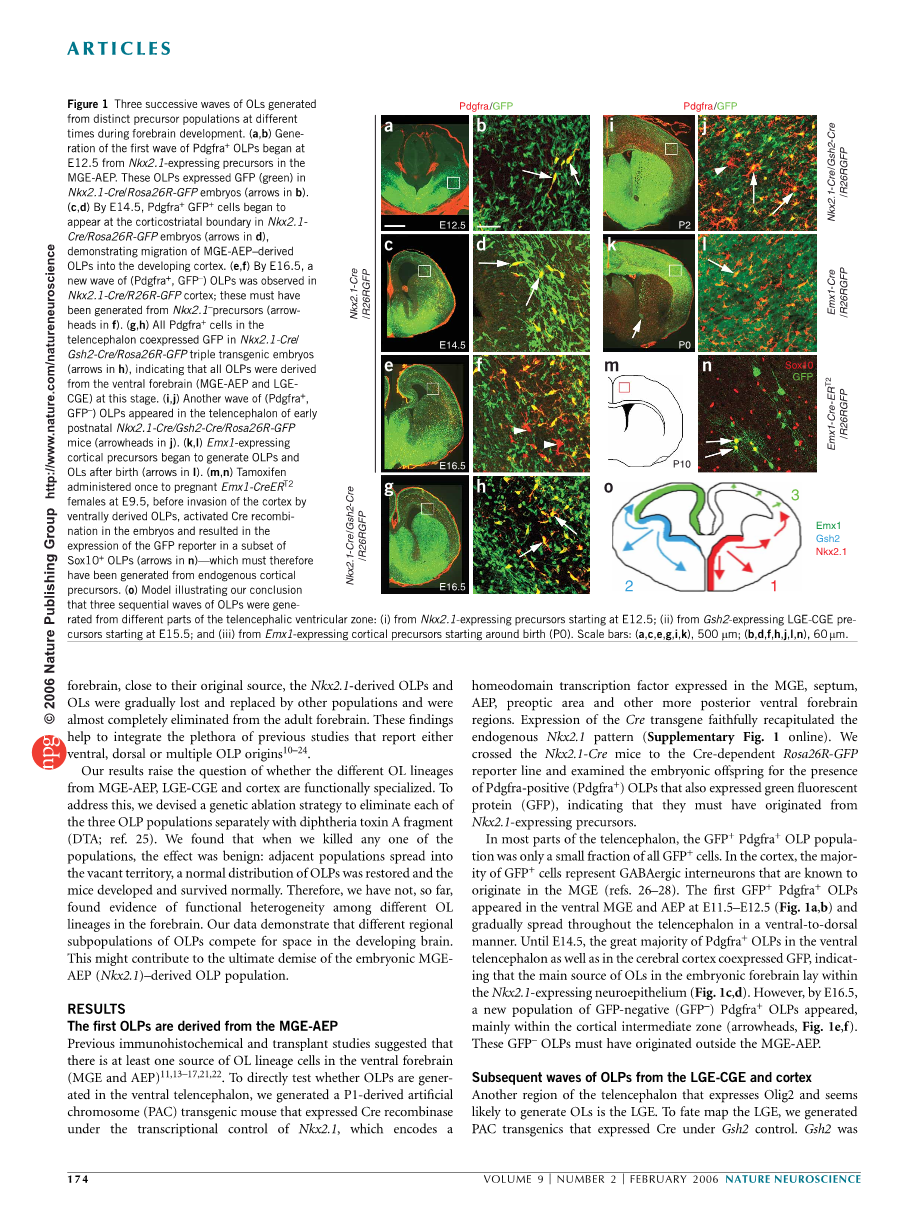
图1 在前脑发育的不同时间从不同前体群体产生的三个连续的OLs波。(a,b)第一波Pdgfra OLPs的基因比从E12.5开始,从MGE-AEP中表达Nkx2.1的前体开始。这些OLP在Nkx2.1-Cre / Rosa26R-GFP胚胎中表达GFP(绿色)(箭头b)。(c,d)通过E14.5,Pdgfra GFP 细胞开始出现在Nkx2.1-Cre / Rosa26R-GFP胚胎的皮质纹状体边界处(d中的箭头),表明MGE-AEP衍生的OLPs迁移到发育中的皮层。(e,f)通过E16.5,在Nkx2.1-Cre / R26R-GFP皮层中观察到新的(Pdgfra ,GFP–)OLPs波;这些必须是由Nkx2.1前体产生的(f中的箭头)。(g,h)端脑中的所有Pdgfra 细胞在Nkx2.1-Cre / Gsh2-Cre / Rosa26R-GFP三重转基因胚胎中共表达绿色荧光蛋白(箭头在h),表明所有OLPs源自腹侧前脑(MGE-AEP和LGE-CGE)。(i,j)在产后早期的Nkx2.1-Cre / Gsh2-Cre / Rosa26R-GFP小鼠的端脑中出现(Pdgfra ,GFP–)OLPs的另一波(j中的箭头)。(k,l)表达Emx1的皮质前体在出生后开始产生OLPs和OLs(箭头在l中)。(m,n)在通过腹侧衍生的OLPs侵入皮层前,在E9.5给予怀孕的Emx1-CreERT2雌性一次他莫昔芬一次,在胚胎中激活Cre重组体,并导致绿色荧光蛋白报道子在Sox10 OLPs(n中的箭头)——因此必须由内源性皮层前体产生。(o)模型说明我们的结论,即从远端脑室区的不同部分产生三个连续的OLPs波:(i)从E12.5开始的表达Nkx2.1的前体; (ii)从E15.5开始的表达Gsh2的LGE-CGE前体; 和(iii)来自表达Emx1的皮层前体在出生时开始(P0)。比例尺:(a,c,e,g,i,k),500微米; (b,d,f,h,j,l,n),60微米。
结果
第一个少突胶质细胞起源于MGE-AEP
早先的免疫组织化学方法和转基因研究显示在腹侧前脑中至少有一个来源的OL谱系细胞(MGE和AEP)11,13–17,21,22。为了直接测试在腹腔端端脑内是否出现了OLPs,我们生成了一种p1-衍生的人工染色体(PAC)转基因小鼠,在Nkx2.1的转录控制下表达了Cre重组酶,该基因编码在MGE、鼻中隔、AEP、前视区和其他后腹前脑区表达同源域转录因子。Cre基因的表达准确地再现了内源性Nkx2.1模式(补充图1在线)。我们穿过Nkx2.1 - Cre小鼠到依赖于Cre的Rosa26R - GFP报告线,并检查了胚胎后代阳性Pdgfra (Pdgfra )的存在,并表达了绿色荧光蛋白(GFP),表明它们一定来自Nkx2.1 -表达前体。
在端脑的大部分区域,GFP Pdgfra OLP种群是所有GFP 细胞的一小部分。在皮质中,GFP 细胞的主要特征是GABAergic间神经元,这是已知的起源于MGE(参考26-28)。第一个GFP Pdgfra OLPs出现于E11.5 - E12.5(图1a,b)的腹腔MGE和AEP中,并逐渐以腹侧到背侧的方式传播到端脑。直到E14.5,腹侧端脑和大脑皮层的绝大多数Pdgfra OLPs和大脑皮层都表达了GFP,这表明胚胎前脑中的OLs主要来源在Nkx2.1表达的神经上皮内(图1c,d)。然而,在E16.5中,新的阴性GFP (GFP –)Pdgfra OLPs出现了,主要在皮质中间区(箭头,图1e,f)。这些GFP –OLPs肯定是来自于MGE-AEP外部。
随后的波来自LGE-CGE和皮质
另一种表达Olig2的端脑区似乎很可能产生OLs的是LGE。对于LGE的命运图,我们生成了在Gsh2控制下表达Cre的PAC 基因转移。Gsh2是在MGE中强烈表达的LGE-CGE,但部分与Nkx2.1重叠(补充图1)。为了绘制腹侧前脑的总体贡献,我们生成了三基因Gsh2 - Cre /Nkx2.1- Cre / Rosa26rR- GFP小鼠。在E16.5三种转基因技术中,端脑中几乎所有的Pdgfra OLPS,包括外侧皮质,也都是GFP (图1g,h)。这表明,在出生之前,大脑皮层的所有OLPs都是由腹侧衍生的,首先从MGE-AEP迁移,然后从LGE、CGE或两者中迁移。然而,在出生的那天,GFP - OLPs再次开始在三重转基因(箭头,图1i,j)的皮层中积累,表明至少有一个额外的来源的OLPs在MGE-AEP或LGE-CGE(可能在皮层本身)之外。
为了检查内源性皮质前体的命运,我们生成了Emx1-Cre基因转移(补充图1)。在这些老鼠中,Cre在皮质前体表达强烈, 且激活的GFP报告者在Emx1-Cre / Rosa26R-GFP老鼠中普遍在产后皮层表达(补充图1)。GFP当前遍布整个细胞的细胞质甚至轴突,使纤维束前脑腹侧的皮层投射GFP标记(箭头,图1 k)。Emx1-衍生的(GFP Pdgfra )OLPs在出生时(图1k,l)中首次出现,并在此后迅速增加。Emx1-衍生的OLPs在腹腔端脑端从未见过。
我们的数据表明,皮质的OLPs是由两个从MGE-AEP和LGE-CGE迁移而来的群体和一个常驻的大脑皮层群体组成的。排除这样一种可能性,即Emx1-衍生的OLPs是由腹侧衍生的细胞在它们迁移到皮质区域后开始表达Emxl,我们生成另一个转基因线即在Emxl控制下表达tamoxifen-inducible形式的Cre重组酶(CreERT2)(补充图1)。我们通过在E9.5中使用他莫西芬诱导短暂的Cre活动,在胶质细胞再生之前。因为它莫西芬在48小时内将被清除,所以在之后到E11.5期间从其他地方迁移到皮质的OLPs不能激活Cre。我们分析了P9中用它莫昔芬治疗的小鼠。CNS中的Pdgfra表达仅限于早期的OLPs,一旦细胞退出细胞周期就会下调并开始分化为OLs(参考29,30)。因此我们对Sox10使用了抗体,它在比Pdgfra稍晚一点的OLPs中开始表达,但在Pdgfra被下调后仍然存在于成熟的OLs中(参考31)。与常规的Cre相比,CreERT2 -mediated的重组效率相对较低,因此在整个皮质区都能观察到良好分离的GFP标记细胞或细胞簇。这些包括Sox10 OLs和OLPs,以及细胞形态的神经元和星形胶质细胞(图1 . 1m,n)。GFP Sox10 细胞组成了所有GFP 细胞的20%,并在皮层的所有部位被观察到(箭头,图1n)。我们的结论是,它们是由内源性皮层前体形成的,而不是来自内迁移的细胞。
图2胚胎Nkx2.1衍生的OL谱系在出生后期迅速消除。(a,b)在Nkx2.1-Cre / Rosa26R-GFP小鼠中,GFP (绿色),Sox10 (红色)双阳性OLPs在P10皮层中已经是少数(a中的箭头),并且其贡献由P80下降到零。(c,d)在P10,几乎所有腹部区域中的所有OLPs(例如隔膜)均来源于Nkx2.1表达的前体(MGE-AEP- 衍生的),(绿色荧光蛋白,Sox10双阳性; c中的黄色核),但即使在这里,它们也被P80替换为其他种群(GFP–)。(e-j)Sox10 OL谱系细胞的三种不同群体的比例贡献表示为每个区域中Sox10 细胞总数的百分比(平均值的标准误

资料编号:[281390],资料为PDF文档或Word文档,PDF文档可免费转换为Word
剩余内容已隐藏,支付完成后下载完整资料
英语原文共 7 页,剩余内容已隐藏,支付完成后下载完整资料
资料编号:[281390],资料为PDF文档或Word文档,PDF文档可免费转换为Word
您需要先支付 30元 才能查看全部内容!立即支付
您可能感兴趣的文章
- 饮用水微生物群:一个全面的时空研究,以监测巴黎供水系统的水质外文翻译资料
- 步进电机控制和摩擦模型对复杂机械系统精确定位的影响外文翻译资料
- 具有温湿度控制的开式阴极PEM燃料电池性能的提升外文翻译资料
- 警报定时系统对驾驶员行为的影响:调查驾驶员信任的差异以及根据警报定时对警报的响应外文翻译资料
- 门禁系统的零知识认证解决方案外文翻译资料
- 车辆废气及室外环境中悬浮微粒中有机磷的含量—-个案研究外文翻译资料
- ZigBee协议对城市风力涡轮机的无线监控: 支持应用软件和传感器模块外文翻译资料
- ZigBee系统在医疗保健中提供位置信息和传感器数据传输的方案外文翻译资料
- 基于PLC的模糊控制器在污水处理系统中的应用外文翻译资料
- 光伏并联最大功率点跟踪系统独立应用程序外文翻译资料


