拟南芥npcc6 / nakr1是一种对于韧皮部功能和根分生组织的维护所必需的韧皮部移动金属结合蛋白
原文作者Hui Tian,a Ivan R. Baxter,b,1 Brett Lahner,c Anke Reinders,a David E. Salt,b,c and John M. Warda,2
SODIUM POTASSIUM ROOT DEFECTIVE1(nakr1;以前被称为npcc6)编码一种在韧皮部伴细胞特异表达的一种可溶性的金属结合蛋白。nakr1-1突变体的表型包括高Na 、K 、Rb ,和叶片中淀粉的积累,根短,开花延迟,蔗糖的长距离输送缩短。基于基因芯片的缺失定位,一个7 bp的缺失在NaKR1的一个外显子上被发现,相当于插入了一个提前终止密码子。突变体的表型是由转化的天然基因或nakr1 GFP(绿色荧光蛋白)和NaKR1 -beta;-葡萄糖醛酸酶融合启动子驱动。NAKR1-GFP在韧皮部移动;它从伴细胞进入筛元件并进入根分生组织的一个以前未发现的共质域。嫁接实验表明,高Na 积累主要是由于叶片中nakr1的功能丧失造成的。这支持了在循环中的韧皮部的作用Na 到根限制叶片中钠的积累。根表型的发病恰逢发芽后NaKR1的表达。nakr1-1短根的表型主要是由于在根分生组织的细胞的分裂率下降,表明了在韧皮部nakr1的表达在根分生组织维持中的作用。
简介
植物维管组织由两个平行的长途运输通道组成:木质部和韧皮部。木质部从根部输送水分和矿质营养元素到枝。韧皮部运输糖和其他代谢物,蛋白质和核糖核酸从源(光合或储存)组织到汇,如根,发育叶片,花,种子,和存储组织。被子植物的韧皮部其主要单位是伴随细胞/筛元素复合物。形成导管的筛元件是高分化的细胞,缺乏许多正常的细胞内的细胞器,如线粒体和细胞核。伴胞与筛管分子通过胞间连丝提供提供代谢支持和合成RNA和蛋白质,在筛管中运输。除韧皮部在运输过程中的基本功能减少库组织的碳,韧皮部在植物中执行各种重要的角色。在韧皮部特异表达的基因已经确定,但是只有相对很少一部分韧皮部表达基因的功能被分析。
经典理论认为,叶片中的韧皮部具有明显的促进钠离子的作用,而在分子遗传研究中没有太多的支持。通过shoot2(sas2 的等位基因 HKT1)突变体中拟南芥钠过度积累的表型,Na 转运蛋白HKT1在韧皮部中的功能被提出。然而,目前的共识是,HKT1的主要功能是从木质部除去Na ,从而限制了叶片中钠的积累。在韧皮部中的钠 转运的程度和机制尚不清楚,并已被确定为目前的研究课题,这对于理解钠离子的耐受性很重要。更多的是已知的钾在韧皮部中的作用。钾,伴随着糖,作为一个在韧皮部的主要渗透调节物质。在蔗糖吸收过程中K 通过AKT2/3外流也可到再极化膜电位。韧皮部的一个主要功能是将光合组织中的固定碳运输到库组织,如生长的叶、根、花和种子等。拟南芥Suc2突变体在蔗糖运输到韧皮部这一过程中有缺陷并且表型明显,包括植株矮小,叶片中淀粉的积累,和花色素苷积累。SUC2表型的严重程度指示伴细胞功能的重要性,但只提供有限的关于韧皮部的其他角色的信息,如在控制开花,K 和Na 转运,和分生组织的功能。
胞间连丝(PD)是韧皮部功能的本质。伴胞和筛管是由专门的不同大小的至少67 kD的PD连接。观察绿色荧光蛋白(GFP)融合蛋白的细胞运动已经在确定共质体结构域(由PD连接的细胞群)。GFP融合36到67 kD在同伴细胞的控制下表达,特异SUC2启动子进入韧皮部并在韧皮部易位,但在根尖不移动,虽然自由GFP(27 kD)是能够通过根尖。这组不同大小27~36kD的PD连接原生韧皮部细胞与根尖细胞。鲜为人知的是,通过胞间连丝或序列特异性蛋白质运输的要求。
在这篇文章中,对SODIUM POTASSIUM ROOT DEFECTIVE1(nakr1;以前被称为npcc6)基因,nakr1-1突变体的表型分析和定位,韧皮部运输,和nakr1蛋白金属结合性能进行了介绍。结果为在韧皮部中的钠 再循环提供了证据,在根分生组织中韧皮部的功能,和在根尖未被发现的共质体域。
结果
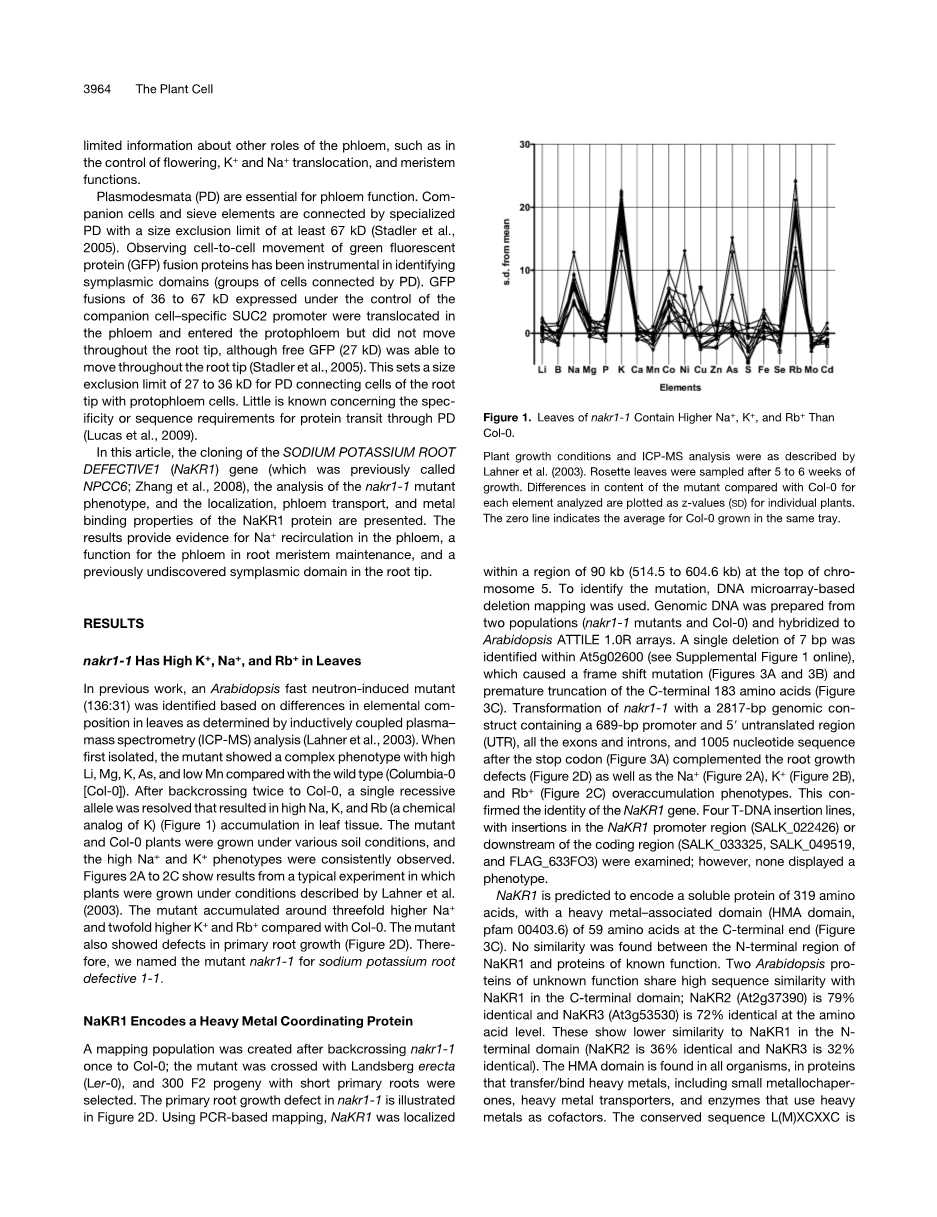
nakr1-1在叶片具有高K ,Na 和Rb
在以往的工作中,快中子诱导拟南芥突变体(136:31)基于叶片中元素组成的差异被鉴定用电感耦合等离子体质谱(ICP-MS)–测定分析。当第一次分离,该突变体表现出复杂的表型与高Li,Mg,K,和与野生型相比较低的Mn (Columbia-0[Col-0])。回交两次Col-0之后,单隐性等位基因得到解决,这导致了叶片组织中高的Na,K,Rb K化学模拟)(图1)积累。这株Col-0植株在不同土壤条件下生长,并持续观察高Na ,K 表型。图2A至图2C示出一个典型的实验结果,这些植株在Lahner等人所描述的条件下生长。与Col-0相比较累积突变在约三倍高Na ,双重高钾,和高Rb 。该突变体还显示初生根生长缺陷(图2)。因此,我们将突变nakr1-1命名为钠钾根缺陷1-1。
nakr1编码一种重金属协调蛋白
在nakr1-1和Col-0回交一次后创建作图群体;突变体与兰兹贝格(Ler-0)杂交,300个短根后代被选择。nakr1-1初生根的生长缺陷在图2D图解。基于PCR的映射,nakr1被定位在5号染色体的顶端90 kb的区域内(514.5~604.6 kb)。为了识别突变,利用基因芯片技术进行基因缺失定位。从2个种群中制备基因组DNA(nakr1-1突变体和Col-0)并与拟南芥ATTILE 1.0R arrays.杂交。一个7 bp缺失被确定在At5g02600(见补充图1),这个缺失导致移码突变(图3a和3b)和C-末端183个氨基酸的过早截断(Figure 3C).用含有2817个碱基的基因组结构一个689 bp的启动子和59个非编码区(UTR)改造nakr1-1,所有外显子和内含子,终止密码子后的1005个核苷酸序列(图3A)补充根系生长缺陷(图2)以及Na (图2a),K (图2b),和Rb (图2C)过度积累的表型。这也证实了NaKR1基因的身份。在nakr1启动子区(SALK_022426)或编码区的下游(SALK_033325, SALK_049519,和FLAG_633FO3)插入四个T-DNA插入线并进行了检查;然而,没有表现出的表型。
图1。与Col-0相比nakr1-1叶片中含有较高的Na ,K ,和Rb 。
植物生长条件和ICP-MS分析如Lahner等人所描述。5至6周的生长后对莲座叶进行取样。分析绘制与Col-0相比突变体的每种元素含量差异的植物个体的z值(SD)。零线表示在同一个托盘中生长Col-0的平均值。
Akr1是预测编码319个氨基酸的蛋白质,在C-末端有一个重金属-相关联的59个氨基酸的域(HMA域,Pfam 403.6)(图3c)。对nakr1和已知功能的蛋白的N末端区域之间没有发现相似性。未知功能的2个拟南芥蛋白分享在nakr1C-末端结构域的高度序列相似性;在氨基酸水平NaKR2 (At2g37390) 79%是相同的,NaKR3 (At3g53530) 72%是相同的,这些与NaKR1 在N末端结构域显示较低的相似性(NaKR2是36%相同,NaKR3 32%相同)。HMA域被发现在所有的生物中,在蛋白质中它转移/结合重金属,包括小的metallochaper,重金属转运蛋白,和使用重金属作为辅助因子的酶。保守序列L(M)xcxxc被认为是一个铜结合位点,其中两个Cys残基直接协调金属,这个结构域也运输蛋白,专门运输Zn2 、Ni 、Hg 。
NaKR1的HMA域分别和拟南芥铜伴侣ATX1 和 CCH共有47 和 45%,同一性。在酵母中的同源物(酿酒酵母)ATX1已经有很好的表征,在通过专门向重金属P型ATP酶提供铜来保持铜平衡的酶和防御氧化应激中都有发现。当酵母菌株中过度缺乏超氧化物歧化酶基因SOD1,AtATX1 和 At-CCH都可以以铜依赖的方式保护突变体的活性氧毒性。在我们的实验中,对nakr1或C-端区域单独表达未能补充酵母sod1d突变体的活性氧毒性表型(见补充图2),表明在nakr1中HMA域的功能不同于At-ATX1/At-CCH在酵母中的表达。
为了测试可能的金属结合,NaKR1作为融合麦芽糖-结合蛋白质在大肠杆菌中表达(MBP)。MBP-NaKR1融合蛋白的纯化在图4a显示。纯化的融合蛋白的MBP-NaKR1身份经质谱分析证实。对四十个独特的肽进行检测,覆盖了融合蛋白序列的48.5%(见补充图3)。在纯化的蛋白质样品中通过ICP-MS分析检测到几种重金属(图4B)。mbp-nakr1融合蛋白与MBP控制相比较有较高的Zn、Cu、Fe、Ni、CO。蛋白质与金属的摩尔比为3:1左右(图4c),表明nakr1是金属结合蛋白。变异版本的nakr1全基因构建在HMA域的两个保守的半胱氨酸残基上,这两个保守的Cys残基转变为甘氨酸,使得未能补充nakr1-1短根和晚花表型(数据未显示)。这表明,在植物中NaKR1的金属结合功能是必不可少的。
NaKR1在韧皮部特异表达
为了研究表达模式, 一个完整的beta;-葡萄糖醛酸(GUS)酶融合基因(NaKR1pro:NaKR1-GUS)在Col-0中表达,并且分析10个独立的转化。在胚胎或吸胀(但没有发芽)的种子中没有发现GUS染色(图5A)。在发芽后一天,在10%的幼苗中观察到GUS染色。在根茎交界处的脉管系统中发现GUS活性并且较小的程度上在下胚轴(图5b)。在2天后在整个幼苗的脉管组织中发现GUS活性(图5c)。在更为成熟的植物中,强的GUS染色被发现在莲座的维管组(图 5D)织和茎生叶(图5E)中。在花组织,中GUS染色主要定位于萼片和花的花丝和角果的花梗。(图5F和5G)。根和叶的叶柄横截面显示表达在脉管系统的韧皮部,而不是在木质部。
图2。nakr1-1 Na, K, and Rb积累表型和由NaKR1全基因补充构建的根系生长缺陷
(A) 至 (C)芽Na , K 的浓度和Col-0, nakr1-1中Rb 的浓度补充线1和3(T2代)作为平均值plusmn;SD (n= 12). 学生没有进行测试,不同字母显示P lt; 0.05的显著差异植物生长、样品采集和ICP-MS分析根据lahner等人。(2003).
(D) 八日龄幼苗在ATS介质中垂直生长。从左到右分别是两个互补的线条,nakr1-1,与Col-0。注意,nakr1-1减少了初生根的生长。
NaKR1 表达(At5g02600) 此前曾报道称为同伴细胞特异性。这是基于基因芯片结果和cDNA文库对分离纯化的伴侣细胞原生质体,核,或孤立的韧皮部组织的转录本分析的结果。在张等人的研究中(2008), NaKR1(在这项研究中被称为NPCC6)在排名前12同伴细胞核中最丰富的转录本。此外,NaKR1pro:histoneB2-GFP在拟南芥中表达并 特别地显示标记的同伴细胞
在我们的实验中,histoneB2-GFP的表达由根中韧皮部的被NaKR1启动子标记的细胞核驱动。(图 6A),由张等人确认的结果。(2008). 在根尖,histoneb2 GFP荧光仅在韧皮部中发现(图6B)。苯胺蓝染色的筛板鉴定在筛管毗邻细胞核中含histoneb2 GFP荧光(图6C)。这证实了NaKR1是在同伴细胞中表达的。整个基因GFP(NaKR1pro:NaKR1-GFP)融合互补的nakr1-1突变体的表型(见补充图4),这表明NaKR1-GFP融合蛋白的功能。T2代幼苗使用共聚焦显微镜进行观察。在根中,GFP荧光被发现在维管内木质部两侧的平行细胞内指示韧皮部定位(图6D)。在根尖,NaKR1-GFP荧光在原细胞分裂区近端的细胞中观察到(图6E)和横向根(图6F)。在细胞质和细胞核的荧光明显(图6E,插图)。
图3。NaKR1基因和蛋白质的结构。
(A) NaKR1基因结构(用于互补),起始和终止密码子的位置突出。从左到右分别是启动子区和5′UTR(ATG前面的黑块和灰块),三个外显子,两个内含子,和5′UTR,黑色箭头表示nakr1-1突变位置。
(B) 在nakr1-1第一外显子缺失7个碱基,由框出的序列表明,造成移码和C-末端氨基酸序列截断。Mut,突变;WT,野生型。
(C) NaKR1的蛋白质结构有重金属-相关领域的C-末端和与其他功能域有一点相似的N端区域。氨基酸位置137指示突变蛋白中的第一变位。
图4。MBP-NaKR1纯化融合蛋白结合重金属
MBP-NaKR1 或 MBP (对照) 在大肠杆菌BL21-AI中表达并纯化使用直链淀粉
(A) 考马斯亮蓝染色聚丙烯酰胺凝胶(12%)显示3 mg纯化mbp-nakr1(条带2)和3毫克的MBP(条带3)。条带1包含低范围SDS-PAGE标准(Bio-Rad)。
(B) 纯化后的蛋白样品MBP-NaKR1和MBP的金属含量。从2个独立的实验得到
剩余内容已隐藏,支付完成后下载完整资料

英语原文共 27 页,剩余内容已隐藏,支付完成后下载完整资料
资料编号:[286298],资料为PDF文档或Word文档,PDF文档可免费转换为Word
您可能感兴趣的文章
- 饮用水微生物群:一个全面的时空研究,以监测巴黎供水系统的水质外文翻译资料
- 步进电机控制和摩擦模型对复杂机械系统精确定位的影响外文翻译资料
- 具有温湿度控制的开式阴极PEM燃料电池性能的提升外文翻译资料
- 警报定时系统对驾驶员行为的影响:调查驾驶员信任的差异以及根据警报定时对警报的响应外文翻译资料
- 门禁系统的零知识认证解决方案外文翻译资料
- 车辆废气及室外环境中悬浮微粒中有机磷的含量—-个案研究外文翻译资料
- ZigBee协议对城市风力涡轮机的无线监控: 支持应用软件和传感器模块外文翻译资料
- ZigBee系统在医疗保健中提供位置信息和传感器数据传输的方案外文翻译资料
- 基于PLC的模糊控制器在污水处理系统中的应用外文翻译资料
- 光伏并联最大功率点跟踪系统独立应用程序外文翻译资料


