英国成熟野樱的快速繁殖
原文作者 N. Hammatt amp; N.J. Grant
摘要:从英国林地野樱(Prunus avium L.)取得的茎尖,也是P. avium的砧木。F12/ 1和外植体成功建立了体外培养,大部分都容易在MS为基础的培养基上进行快速繁殖。在一种途径,不定芽偶尔由位于初始外植体的叶柄的花外蜜腺发育而来。快繁简介:外植体用Quoirin和Lepoivre还有以植物为基础的木本植物培养基来改善,并通过用1-苯基-3-(1-,2-,3-噻二唑-5-基)脲(噻苯隆)补充培养基。丛生芽按F12/1无1,3,5三羟基苯(间苯三酚)中心结构的生产在最多至18个月定期传代培养中不改善。研究简介: F12/ 1表明,用1mM间苯三酚,0.49mu;M吲哚丁酸,4.4mu;M苄基腺嘌呤(BA)和0.29mu;M赤霉酸(GA3)一个基于MS的培养基上产生的枝条,比较容易在几个传代培养生根超过在无赤霉素类似介质和仅2.2mu;M BA的培养基上生芽,但只有在生根培养基补充有1mM的间苯三酚时。
关键词: 克隆,农场的林地,林业,繁殖,Prunus avium L.,再生。
引言
野樱桃(P.avium)是一个快速成长的,高价值,耐松鼠,硬木树(普赖尔,1985年,1988年),具有为林业和农业林地广泛应用的潜力。迄今为止,用推荐的做法在英国种植野樱的种子通常是从未经检验的且是进口的种子。这种做法已导致在预测生产力和所得到的林地经济可行性是有困难。这些问题可能通过种植克隆的良好的性能的树木被规避。从长期来看,基因特点,可以从育种计划(尼科尔,1993年)预期改进种植材料。同时,高于平均水平的克隆可以派生自现有的林地中选择的树木,以及可能从CVS。如F12/ 1和外植体,其被开发用于作为振兴砧木甜樱桃的生产,并具有抵抗由Pseudomonas syringae pv. morsprunorum(加勒特,1988)引起的细菌性溃疡病的能力。
利用根(Ghani和Cahalan,1991年),和芽(A1 Barazi和Schwabe,1985)切段成功克隆了P.avium的报道很少。进一步来说,一般情况,当树木成熟和上了年龄,从它们上获取的切段生根越来越困难。快繁,可一年四季进行,可以提供另一种选择,迅速的方式产生野樱克隆。这些实验的目的之一是发展来自英国林地成年野樱桃树使用的快繁方案,也是砧木F12/ 1和外植体。另一个目的是评估用于生产芽的BA浓度是否会影响他们以后形成根的能力。
材料和方法
1.植物材料
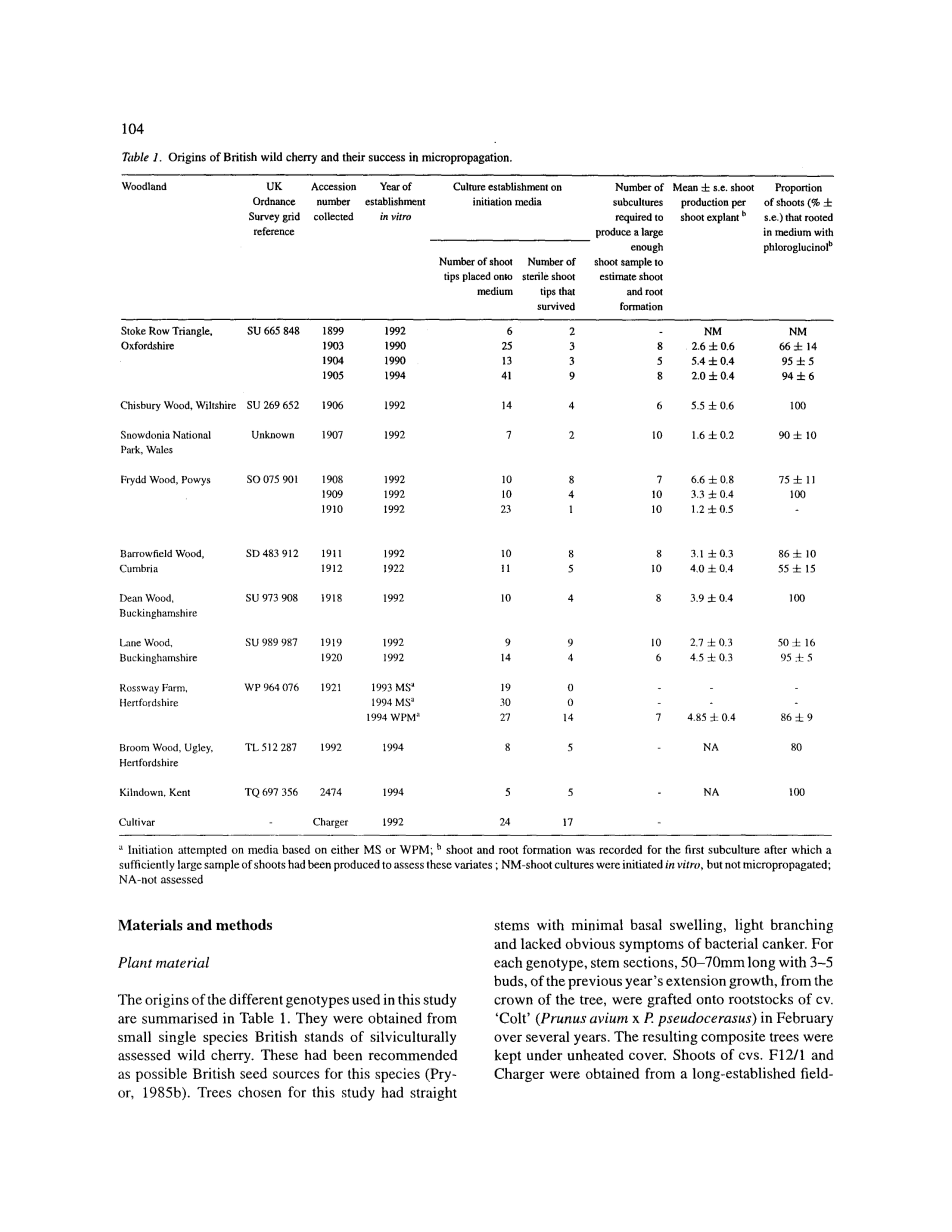
在这项研究中所用的不同基因型的起源总结于表1中。他们从小型单一物种的英国人工林里评估的林营野生樱桃获得的。这些已被推荐为这个物种的可能是英国的种源(普赖尔,1985年b)。本研究选择了树直具有最小基底肿胀,少分枝和缺乏细菌性溃疡病的明显症状。对于每个基因型,前一年的扩大生长的切段末端,从树的树冠,有50-70mm长的3-5个芽,被嫁接到栽培变种的砧木上。雄(Prunus avium x P. pseudocerasus)在数年内的二月,由此产生的复合树木被控制在未加热掩护下保存。 突变种的芽。从一个历史悠久的田间或得的F12/ 1和外植体,也嫁接到“雄”(1980年),并每年春天一直定期修剪。
2.培养基
大多数的培养基基于矿物盐和Murashige和Skoog(1962),指定的MS的有机成分,但是用30mu;m的ZnS04.7H20和100mu;m的FeNaEDTA代替ZnSO4.4H20,NA2-EDTA和FeS04.7H2O,和没有水解物,吲哚乙酸或激动素。其他培养基是基于木本植物培养基的A-H(WPM; Lloyd and McCown,1981)化学成分或Quoirin和Lepoivre(1977)的盐与WPM的组分G和 H。后者中被命名为QL/ WP。所有的生芽和生根培养基含有87.6mM蔗糖和6.0gl-l西格玛琼脂(pH值=5.6)。
3.培养物建立
茎尖采取自在五月和六月迅速伸出的嫩枝,表面消毒通过浸在10%(V / V)商业消毒剂“Domestos”(利华兄弟公司,伦敦;0.8%w / v的终浓度次氯酸钠)中10分钟,随后用无菌水彻底冲洗。每个芽最上面5毫米被切除,及其所有叶原基10mm以上被切除。所得外植体放入30毫升由一层聚苯乙烯主体和聚乙烯帽(格雷纳实验室技术,德思礼,格罗斯,UK)组成的库尔特计数器杯(图1)。每杯含8毫升各种起始培养基。培养物是通过每隔一个月转移到各种新鲜的芽培养基进行传代。
4.生根
一旦幼芽足够每个克隆(以实现每个实验的足够嫩枝需要传代培养的数目列于表1中所示),将对它们的生根能力进行测试,通过将枝条在生芽培养基继代培养28天后,转入装有8毫升生根培养基的柯尔特计数器杯子。新的培养基由MS和14.7mu;M IBA组成,不论有无1mM的间苯三酚(每杯两杆)。芽生根的比例在下一个28天之后计算。
5.外植体的快繁
在含有1 mM间苯三酚的基础MS培养基中经过16个继代周期,2.2 mu;m BA和 0.49 mu;M IBA,外植体的芽被区分和保持在4个继代周期中继代与无论是MS或QL/WP基础培养基。都被具有或不具有50 nM TDZ测试过。
6.预处理 F12/1的芽来增加在有间苯三酚的培养基的生根率
对于F12/1,一旦有4个腋芽产生于两条建立于包含4.4mu;M BA、0.49mu;M IBA和0.29Mu;m GA3的MS起始培养基的培养链,但是没有间苯三酚,每个被用于在四种芽增值培养(I-IV)基上建立独立的亚系。这四种培养基分别为起始培养基包含(I)或不包含(II)间苯三酚,以及在起始培养基基础上减少BA为2.2mu;M和移除GA3的包含(III)或不包含(IV)间苯三酚。
7.培养条件
培养物保持在24.6plusmn;2°C,同时每天通过安装在培养物上方24cm处的欧司朗125W白色荧光灯供给90-100mu;mol m-2s-1的光照。
8.移栽生根芽到土里
在转移到生根培养基28天后,每个生根的芽都会被移栽到200 cm3的由腐殖土与纱石为基础的SHL专业全能肥料(威廉·辛克莱,林肯,英国)中。植物被维护在一个370 mmⅹ560 mm的带有透明通风的聚丙烯盖子的聚乙烯植物繁殖托盘(托盘加盖子共225 mm高)。在7天的封闭培养后,逐步打开通风口,指导植物在外界环境中不再枯萎(转移到土后10-14天)。生根的植物被维持在24°C且在植物的上方由飞利浦的36W/84日光灯管给予每天16小时的150 mu;mol m-2s-1的光照,同时维持70%的相对湿度。
9.统计分析
“外植体”和F12/1的数据分析是用逻辑回归技术(斯内德克和柯克伦,1980)。对于“外植体”,每4个重复实验中20个芽被置于一个培养基中且是完全随机的。对于F12/1,平均芽数和生根能力从每个亚种的不同芽数计算得到,但是这个芽数不超过24,生根能力不超过20。这种变化是由于传代培养的不足以及偶尔的污染损失造成的。因此分析差异的时候,通过利用GENSTAT V(Genstat 5 Committee 1987)程序来分析F12/1芽和根的数量,芽样本中在大小方面被加权了。
结果
1.起始培养
所有林质种质的接穗都成功嫁接到了“雄”砧木上。小于10%的初始茎尖外植体在任何克隆中有明显的微生物污染,后者被丢弃。F12/1、“外植体”和所有的木质克隆的茎尖,出来登记的1921年,确定的生芽培养基为在MS为基础的培养基中加入4.4 mu;M BA、0.49 mu;M IBA、0.29 mu;M GA3和1 mM间苯三酚;1921年的茎尖只确定为WPM基础培养基加入4.4 mu;M BA、0.49 mu;M IBA、0.29 mu;M GA3和1 mM间苯三酚。存活并被转入生根培养基的未被污染的茎尖都统计在表1中。在MS基础培养基中四个星期后,可以推测出1906年的克隆中不定芽是从位于叶柄远策外蜜腺发出的(图二)。
2.林地基因型快繁
在林地基因型的芽开始在起始培养基上增殖的时候,它们被转入以及维持在生芽培养基上[有2.2 mu;M BA、0.49 mu;M IBA和1mM间三苯酚(Hammatt和Grant, 1993)的MS培养基]。
在这个培养基上,在每一个连续培养期内,除了1907和1910的所有基因型平均都发展了至少一种除了原有外植体(表1)的茎尖的腋芽.在所有情况中,平均芽数在每一培养周期中不受连续培养的影响。
任何基因型的芽在含有2.2 mu;M BA、0.49 mu;M IBA和1mM间三苯酚的MS培养基上都不是从花外蜜中生出的。1906的不定芽在起始培养基上都发生于蜜腺,在转入有2.2 mu;M BA、0.49 mu;M IBA和1mM间三苯酚的MS基础培养基后发展更好,同时伸长及发生腋芽的方式与初始外植体茎尖发出腋芽的方式相似。
所有被测基因型的生根(图一)在无论是有或没有间三苯酚的培养基中都实现了。有间三苯酚的培养基的实验结果在表一。生根的幼苗全都成功地在SHL堆肥中生长建立。为了测试利用这些生根及生芽培养基的繁殖速度潜能,随机挑选了建立于1994年5月的编号为2474的茎尖,在初始培养的4个月内产生了68个生根芽。
3.“外植体”的快繁
总的来说,在实验(plt;0.001)中芽数有显著的变化;汇集收集到的关于4个实验的数据显示,QL/WP(plt;0.01)和TDZ(plt;0.001)的芽数增长都比没有TDZ(表二)的MS培养基要显著。“外植体”的芽都成功地在SHL堆肥中生根且建立。
4.为了提高F12/1在有间三苯酚的培养基中的生根率进行预处理
从来自在芽增值和生根培养基上的两个独立茎尖的培养系数据整合分析来看,两者的表现没有明显的差异。芽数不受BA浓度(图三)影响,同时在有或无间三苯酚的培养基中连续继代培养没有明显的增长。然而,芽数整体是显著(plt;0.001)提高的通过添加间三苯酚。当在有2.2 mu;M BA、0.49 mu;M IBA,没有GA3和间苯三酚的培养基中继代了480天后,转入有1mM间苯三酚的相似培养基中后,芽数马上提升了。在两个传代培养的芽数变得和在后来的培养基上的亚系刚建立的开始后培养的建立时。
用于生根的芽是来自维持在含有间三苯酚的芽增殖培养基的芽繁殖链。整体来说,通过往生根培养基中添加间三苯酚可以提高生根率(plt;0.001;图四)。用于生产芽的BA浓度和用于生根培养基的间三苯酚,两者对于重复继代的生根效果有一个明显的增长(plt;0.01)。关于处理效果更详细的分析表明连续继代芽生根的比例更高(plt;0.001;图四),但是只有在没有间三苯酚的生根培养基中。另外,在有4.4 mu;M BA、0.49 mu;M IBA和0.29 mu;M GA3及间三苯酚的MS生根培养基中比有2.2 mu;M BA和0.49 mu;M IBA(图四a,b)有更高的芽数(plt;0.05)。在只有IBA的培养基(图四a,b)中,BA浓度以及GA3的浓度对生根效果没有影响。
结论
相比较之前的只设计少量P.avium基因型(例如Snir, 1982; Biondi et al., 1990, Ochatt, 1991)的研究,目前的工作证明了快繁模式在已经报道的品种的成功。F12/1(Hammatt和 Grant, 1993)可以被用于克隆大范围的成熟的林地野樱树。另外,克隆2474的结果表明快繁可以提供比通过芽或根扦插的受季节影响的传统繁殖方式更快捷的培养数的途径。
剩余内容已隐藏,支付完成后下载完整资料

英语原文共 8 页,剩余内容已隐藏,支付完成后下载完整资料
资料编号:[286292],资料为PDF文档或Word文档,PDF文档可免费转换为Word
您可能感兴趣的文章
- 饮用水微生物群:一个全面的时空研究,以监测巴黎供水系统的水质外文翻译资料
- 步进电机控制和摩擦模型对复杂机械系统精确定位的影响外文翻译资料
- 具有温湿度控制的开式阴极PEM燃料电池性能的提升外文翻译资料
- 警报定时系统对驾驶员行为的影响:调查驾驶员信任的差异以及根据警报定时对警报的响应外文翻译资料
- 门禁系统的零知识认证解决方案外文翻译资料
- 车辆废气及室外环境中悬浮微粒中有机磷的含量—-个案研究外文翻译资料
- ZigBee协议对城市风力涡轮机的无线监控: 支持应用软件和传感器模块外文翻译资料
- ZigBee系统在医疗保健中提供位置信息和传感器数据传输的方案外文翻译资料
- 基于PLC的模糊控制器在污水处理系统中的应用外文翻译资料
- 光伏并联最大功率点跟踪系统独立应用程序外文翻译资料


